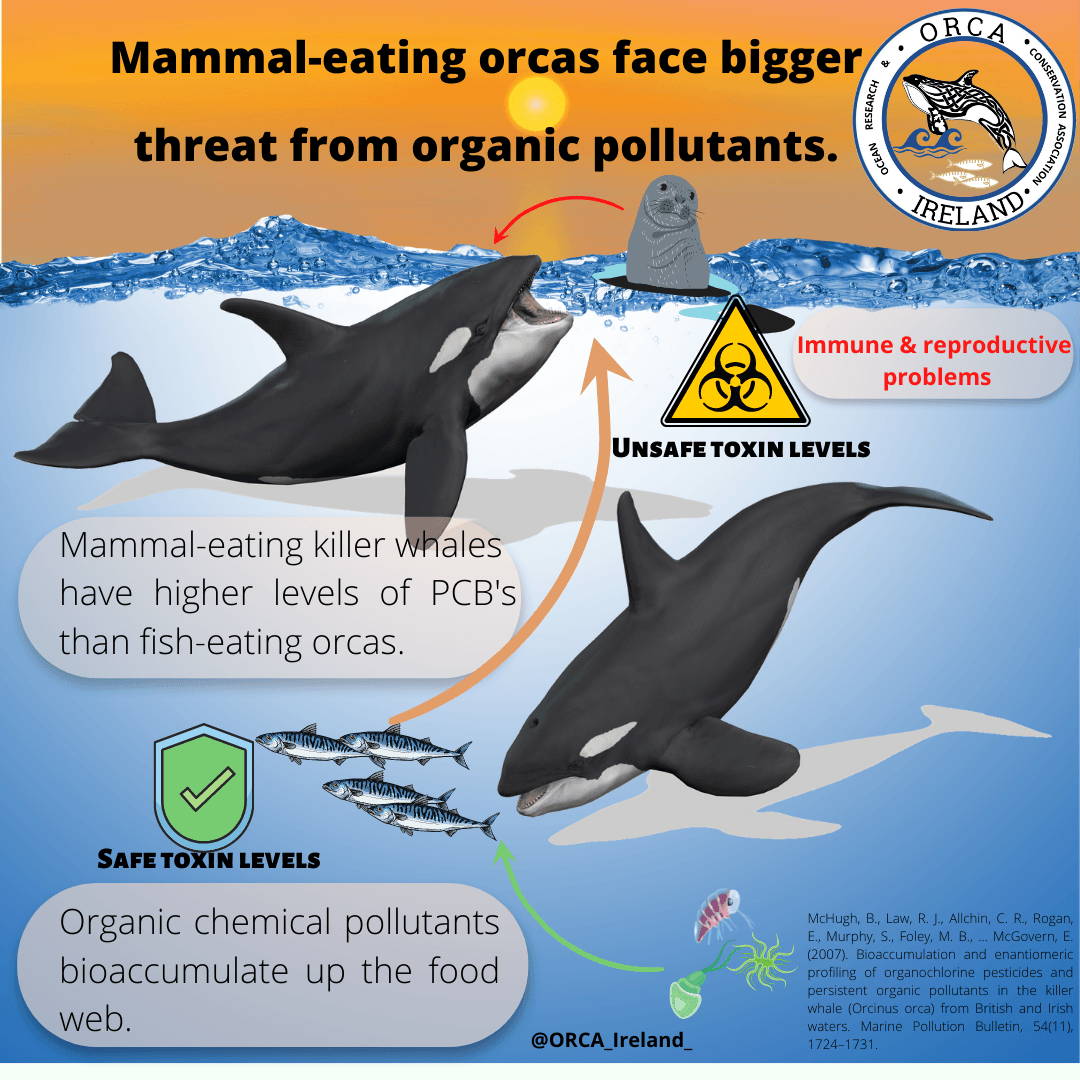
Deep-diving Adaptations in the Sperm Whale.
The sperm whale is one of the deepest diving cetaceans and typically undertake dives for 45 minutes to depths of more than 1,000 meters. They conduct these deep dives to forage on their primary prey, the giant squid. But how are they able to go this deep? How do they manage to overcome the challenges of the deep sea?
The deep sea is considered a hostile habitat, with its high pressure and low levels of oxygen. For a positively buoyant species like the sperm whale, even just getting to the deep is a challenge because naturally its body wants to stay close to the surface. To overcome these challenges, and travel to the location of their prey, sperm whales have evolved several different adaptations which enables this behaviour.
Sperm whales (Physeter macrocephalus) have a large and streamlined body, which reduces drag and energy spent during dives (Reeves & Stuart, 2003). The large body size makes for a great oxygen storage, which means that the whale can obtain aerobic metabolism during deep dives. The sperm whale also has a high concentration of myoglobin in its skeletal muscle. Myoglobin is an oxygen-storing protein, and further facilitates aerobic metabolism (Noren & Williams, 2000). When the oxygen storage is empty, the whale has reached its aerobic dive limit (ADL), and metabolism continues anaerobically. Anaerobic metabolism is less efficient and produces lactic acid which negatively affects locomotion. Most voluntary dives therefore stay within the ADL (Kooyman et al. , 1981).

After submersion, there is a reduced distribution of blood to organs and tissue, making them anaerobic. This may also lower metabolism. The oxygenated blood is redistributed to the heart and brain, which are essential organs when the whale is faced with asphyxia (condition of deficient supply of oxygen). This process is known as peripheral vasoconstriction (Elsner and Gooden, 1983). Odontocetes rely on peripheral vasoconstriction along with bradycardia, for supply of oxygen to the heart and brain during deep dives (Kooyman et al ., 1981).
To extend breath hold capacity, the sperm whale lowers its heart rate and cardiac output, this is known as bradycardia (Kooyman et al., 1981). The blood-flow from the heart is directed towards organs with little anaerobic capacity, such as the brain. In addition, they have vascular nets called retia mirabilia adjacent to the brain, which serves as oxygen storages. These nets are crucial during deep prolonged dives, as they supply the brain with oxygenated blood. The retia mirabilia might also be able to trap air bubbles, and thereby preventing whales from getting “the bends”, which is a possibly fatal decompression sickness (Bonato et al ., 2019).
The sperm whale has an organ inside its head, filled with spermaceti oil. This oil helps regulate buoyancy and enables the whale to glide down to the deep. During a dive, the oil is cooled down from around 33°C to 30°C. It is not yet known exactly how this happens; however, one suggestion is that cool water enters the nasal passages and cools down the oil. When oil freezes, the whale’s density increases, it becomes neutrally buoyant and glides more easily towards the deep. During the ascent, this process is reversed, and the whale become positively buoyant again (Clarke, 1978).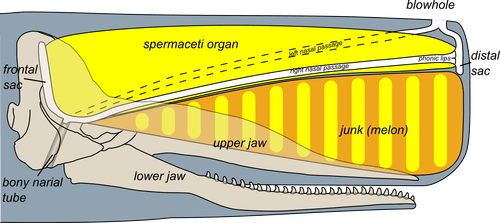
Water pressure increases about one atmosphere per 10 meters in seawater, which means that at 2000 meters depth, which is normal for a sperm whale dive, the pressure is approximately 200 times the pressure at the surface. To deal with this enormous pressure, P. macrocephalus has elastic lungs and a stiff trachea. During dives, the lungs compress and forces oxygen and nitrogen into the trachea, where it is stored until the whale ascends again. This adaptation prevents gases from entering the blood and creating air bubbles (the bends). In addition, the whale has flexible ribcages which allows for expansion and compression of the lungs (Bostrom et al., 2008; Leith, 1989).
Many of these adaptations are also seen in other deep diving cetaceans and are therefore not limited to the sperm whale. All these adaptations together enable the sperm whale to reach the depths of which it can find its favourite prey, the giant squid.
References :
Watwood, S.L., Miller, P.J.O., Johnson, M., Madsen, P.T., Tyack, P.L., (2006), Deep-diving foraging behaviour of sperm whales ( Physeter macrocephalus ) , Journal of Animal Ecology, Vol. 75, pages 814-825
Noren, S.R., Williams, T.M., (2000), Body size and skeletal muscle myoglobin of cetaceans: adaptations for maximizing dive duration, Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, Vol. 126, Issue 2, pages 181-191
Kooyman, G.L., Castellini, M.A., Davis, R.W., (1981), Physiology of diving in marine mammals, Annual Reviews Physiology, Vol. 43, pages 343-356
Bonato, M., Bagnoli, P., Centelleghe, C., Maric, M., Brocca, G., Mazzariol, S., Cozzi, B., (2019), Dynamics of blood circulation during diving in the bottlenose dolphin (Tursiops truncates): the role of retia mirabilia, Journal of Experimental Biology, DOI: 10.1242/jeb.198457
Clarke, M.R., (1978), Buoyancy control as a function of the spermaceti organ in the sperm whale, Journal of Marine Biology, Vol. 58, pages 27-71
Bostrom, B.L., Fahlman, A., Jones, D.R., (2008), Tracheal compression delays alveolar collapse during deep diving in marine mammals, Respiratory Physiology and Neurobiology, Vol. 161, Issue 3, pages 298-305
Leith, D.E., (1989), Adaptations to deep breath-hold diving: respiratory and circulatory mechanics, Undersea Biomedical Research, Vol. 16, Issue 5, pages 345-354
SHARE THIS ARTICLE